
lucille
- balanced physique
- compound movements
- April 26, 2024
- 0 Comments
Master the Moves Full Body Compound Exercises List
Read more about full body compound exercises list
…

Health

Health
7
58
120
52
9
55
11
42
8
278
3
113
86
11
87
106
47
12
10
135
120
69
133
4
66
67
44
59

Dynamic Circuit Bliss for Full-Body Transformation
Embarking on a fitness journey is like diving into a realm of possibilities, and when it comes to transforming your physique, full-body circuit training stands out as the beacon of dynamic bliss. It’s not just about sweating it out; it’s about sculpting a masterpiece, one circuit at a time.
Ignite Your Workout: Intense Full-Body Circuits
Say goodbye to lackluster workouts that leave you yearning for more. Intense full-body circuits are the ignition your fitness routine craves. Each circuit sets your muscles ablaze, pushing your limits and propelling you towards a level of intensity that

Ultimate Mass Builder: Unleash Your Inner Beast with Full-Body Bulk Workout
Are you tired of the same old gym routine that only seems to target specific muscle groups? It’s time to break free from the monotony and embark on a journey of total body transformation. Get ready to unleash your inner beast with the ultimate full-body bulk workout that promises to redefine your physique.
Total Body Blitz: The Wholeness Workout
Say goodbye to isolated exercises and hello to the Wholeness Workout – a comprehensive routine that engages every muscle group in your body. This total body blitz is designed to

Unveiling the Full Body Bodybuilding Workouts on Reddit
Dive into the Reddit Fitness Community
When it comes to exploring full body bodybuilding workouts, Reddit emerges as a goldmine of information and shared experiences. The platform hosts a vibrant and diverse fitness community where individuals from all walks of life come together to share tips, tricks, and personal success stories. If you’re on the lookout for comprehensive insights and real-world advice, Reddit is the place to be.
Discovering the Ultimate Guide
Navigating through the vast sea of information on Reddit may seem daunting, but fear not – there’s an ultimate guide
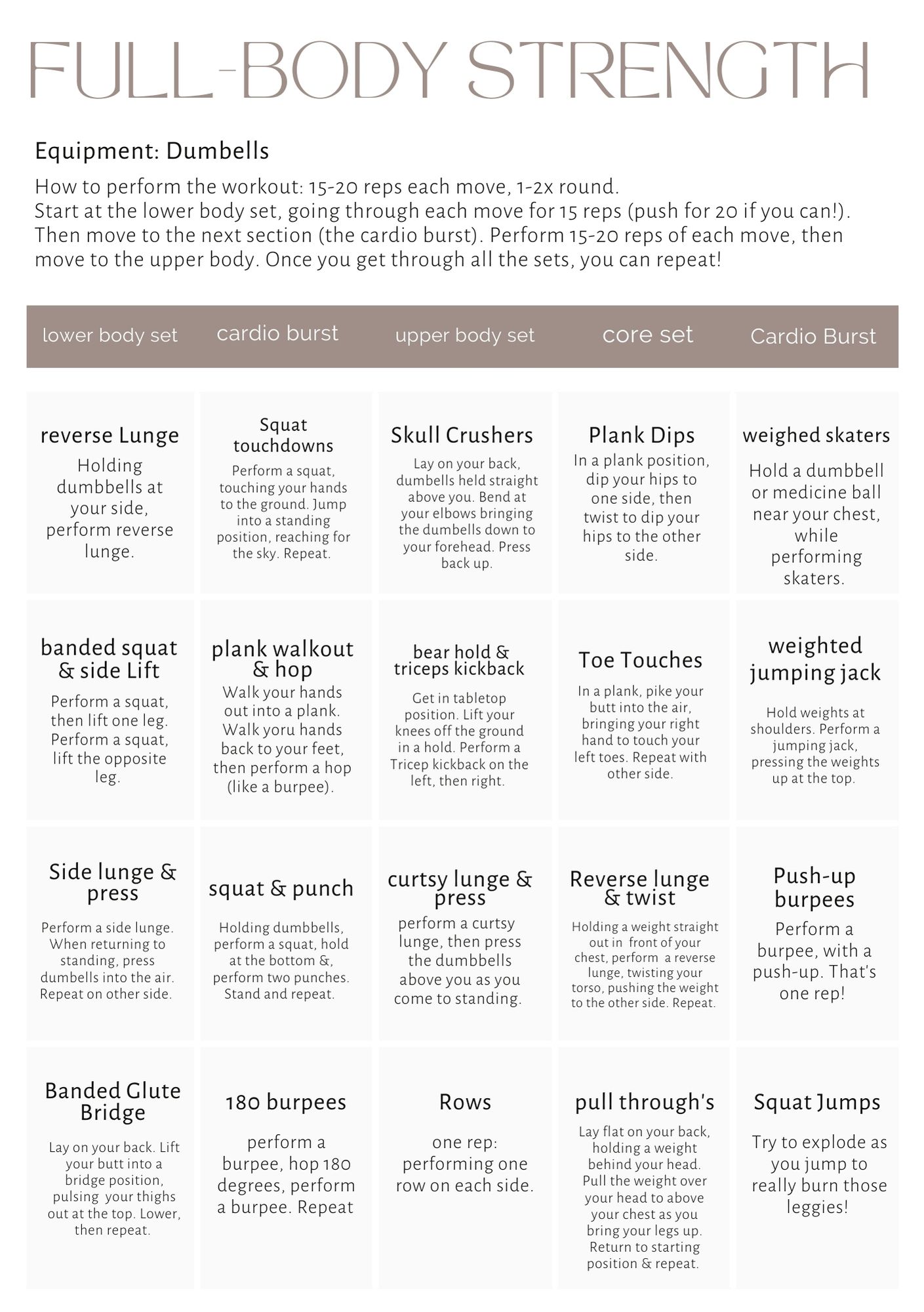
Weighted Bliss: Sculpting Lean Muscles with Full Body Workouts
Are you tired of the same old cardio routine? It’s time to take your workouts to the next level by incorporating weights into your full body cardio sessions. Elevating your routine with weights adds a new dimension to your fitness journey, helping you sculpt lean muscles and achieve a total body transformation.
Full body cardio workouts are already known for their effectiveness in burning calories and improving cardiovascular health. Now, imagine combining the power of cardio with

Unlocking the Power: Full Body Dumbbell Workout for Maximum Muscle Gain
Dumbbells: The Game-Changer for Muscle Growth
In the quest for a formidable physique, the humble dumbbell emerges as a powerhouse. Its simplicity belies its effectiveness in triggering muscle growth. Unlike fixed machines, dumbbells engage stabilizing muscles, promoting a more natural range of motion and ensuring every inch of your body is challenged.
The Essence of Full Body Engagement
Enter the full-body dumbbell workout, a game-changer for those serious about muscle gain. It’s not about isolating specific muscles but engaging your entire body in one cohesive unit. This holistic approach

Sculpting Power: Embark on the Ultimate Full Body Dumbbell Workout Program
Unlocking the Dynamics: Dive into a Comprehensive Routine
Ready to elevate your fitness game? It’s time to unleash the power of a full-body dumbbell workout program. This isn’t just about lifting weights; it’s a dynamic journey that targets every inch of your body. Say goodbye to isolated routines; this program is your ticket to comprehensive gains.
The Core Philosophy: Intense Gains with Dumbbell Mastery
At the heart of this program is the philosophy of intense gains through dumbbell mastery. Dumbbells become your tools for sculpting power, engaging multiple muscle

Dynamic Cardio Blast: Full Body Circuit Unleashed
Get ready to break a sweat and elevate your fitness game with the Dynamic Cardio Blast—a full-body circuit that’s designed to unleash your inner powerhouse. This high-energy workout is not your typical stroll on the treadmill. It’s a symphony of movements that engages every muscle group, leaving you energized, revitalized, and craving more.
The Ultimate Warm-up: Igniting the Cardio Fusion Fiesta
Start your Dynamic Cardio Blast with a bang by incorporating an effective warm-up routine. Engage in dynamic stretches, joint rotations, and light cardio exercises to get your blood pumping and muscles ready

Blaze Your Way to Fitness: Ignite Your Full-Body Burn
Ready to turn up the heat on your fitness journey? It’s time to blaze your way to a fitter, healthier you with the ultimate full-body burn workout. Get ready to torch calories, incinerate fat, and sculpt a physique that radiates strength and confidence.
Embrace the Heat: Unleashing the Total Body Burn Routine
Say goodbye to the mundane and embrace the heat of a total body burn routine. This isn’t your average workout – it’s a fiery experience that targets every inch of your body. From head to toe, each move is

Power Up: Full-Body Circuit Weights Mastery
Embarking on a fitness journey isn’t just about breaking a sweat; it’s about mastering the art of full-body circuit training with weights. It’s time to power up your routine, sculpt your physique, and achieve fitness mastery like never before.
Sculpting Strength: Dynamic Weighted Circuits
Say goodbye to the monotony of traditional workouts and embrace the dynamic power of weighted circuits. Each movement becomes an opportunity to sculpt your strength. It’s not just about lifting; it’s about sculpting and chiseling your body with the transformative power of dynamic, weighted exercises.
Weighted Wonders: Total Body Circuit